Pisa, ottobre 1968
Discorso del Vincitore del Premio Galilei
La genetica delle piante: addomesticamento, miglioramento, genomica
Introduzione. L’occasione concessa allo scrivente con il Premio Galilei ha un importante significato onorifico. Ho ritenuto, per questo, di riassumere nelle Note 1 e 2 il mio curriculum scientifico e gli argomenti di ricerca affrontati in quasi sessanta anni di attività dedicata allo studio delle piante (specialmente di quelle di interesse agrario), attività più o meno direttamente rivolta alla loro genetica e miglioramento genetico, aspetti genomici inclusi. Nel celebrare la sessantesima edizione del Premio Galilei, la Fondazione dei Rotary Italiani intende riassumere in una pubblicazione i testi preparati da coloro che il premio hanno ottenuto; ho aderito all’iniziativa producendo anche un aggiornamento dei temi che furono al tempo (2012) trattati. Tra i variegati interessi scientifici che ho cercato di sviluppare, emergono i due temi che riassumo di seguito: Resistenza alle malattie e Origine delle piante agrarie.
Piante immuni dalle malattie da microrganismi. In agricoltura la riduzione del carico ambientale è correlata in modo diretto alla coltivazione di piante che resistono alle malattie e limitano il ricorso ai trattamenti chimici. Lo sviluppo di queste piante ha considerato una pluralità di strategie e particolarmente i geni R dominanti della famiglia NBS-LRR che, in tutti i vegetali, codifica per i recettori del segnale proveniente dal parassita. Il mio coinvolgimento nell’argomento necessita di una premessa. Quando si cercò di giustificare lo sviluppo di bombe nucleari ricordando a tutti che l’atomo poteva anche produrre cose benefiche, una ragione a favore riguardò l’uso delle radiazioni per indurre mutazioni favorevoli nelle piante agrarie. In preparazione di un libretto Quaderni di Biologia focalizzato sul Miglioramento delle piante agrarie via mutagenesi indotta (Gavazzi e Salamini, 1980), mi incuriosì constatare che le mutazioni per resistenza dell’orzo a tutte le razze di oidio (almeno 11 indipendenti) erano recessive e dovute ad alleli dello stesso gene Mlo. Ne scrissi nel libretto, ripromettendomi di considerare nel futuro questo gene. Attivo a Colonia in un Istituto Max Planck, decisi di attivare la ricerca sul gene Mlo con una mia phD e con un postdoc, Paul Schulze-Lefert: dopo il dottorato in biochimica in un altro dipartimento, questi mi chiese di poter lavorare da noi su questa strana classe di mutanti. Sono stati necessari 10 anni per clonare Mlo: nel 1997 due pubblicazioni riportarono finalmente la localizzazione di Mlo e il suo clonaggio (Hinze et al., 1991; Buschiges et al., 1997; Simons et al., 1997; Wolter et al., 1993). Studi successivi, condotti specialmente sotto la guida di Paul Schulze-Lefert, resero chiaro che Mlo era un gene della pianta necessario alla stessa per qualche ragione, ma che la proteina prodotta era anche utilizzata dal parassita il cui genoma non possedeva quella funzione necessaria per la virulenza. L’approccio basato sulla perdita di funzioni di suscettibilità della pianta sostenute dai geni S (Wang et al., 2018) è ora noto con l’acronimo LOS (Loss of susceptibility), una forma di resistenza ereditata in modo recessivo. L’immunità LOS conferisce una resistenza durevole. A conferma del ruolo futuro dei mutanti recessivi dei geni S nel controllo delle malattie delle piante coltivate, nella Tabella 2 di Zaidi et al. (2018) vengono elencati 19 geni S di 7 specie di interesse agrario, per i quali sono stati indotti, in un breve periodo temporale, mutanti recessivi con metodi di trasformazione genetica o di genome editing. Le resistenze sostenute da questi mutanti riguardano Oidio, Pseudomonas, Xanthomonas, Phytophtora, Virus vari, Colpo di fuoco batterico e altri patogeni. Sono stati isolati altri geni S diversi da Mlo, ma che comunque hanno alleli recessivi responsabili della resistenza. La generalizzazione di questi esperimenti e dei loro risultati ha permesso di sviluppare il concetto di immunità delle piante agrarie alle malattie, basata non più sui geni R (sempre scavalcati da mutazioni del parassita) ma sui geni S. Sono già disponibili, sull’argomento, molte reviews; inoltre, in tutti i genomi vegetali decodificati, i geni Mlo sono sempre presenti in un numero relativamente elevato di copie: il silenziamento in una specie di uno o più di questi geni risulta nella resistenza (il caso di vite e melo; è un esempio italiano; Malnoy et al., 2016; Pessina et al., 2016). È evidente il contributo del nuovo approccio alla sostenibilità della pratica agricola in termini di riduzione dell’uso di prodotti agrochimici contro i parassiti.
Tabella. Mutanti di geni S delle piante. Le citazioni aggiunte nell’ultima colonna della tabella sono citate per esteso in Pozzi e Salamini (2019).
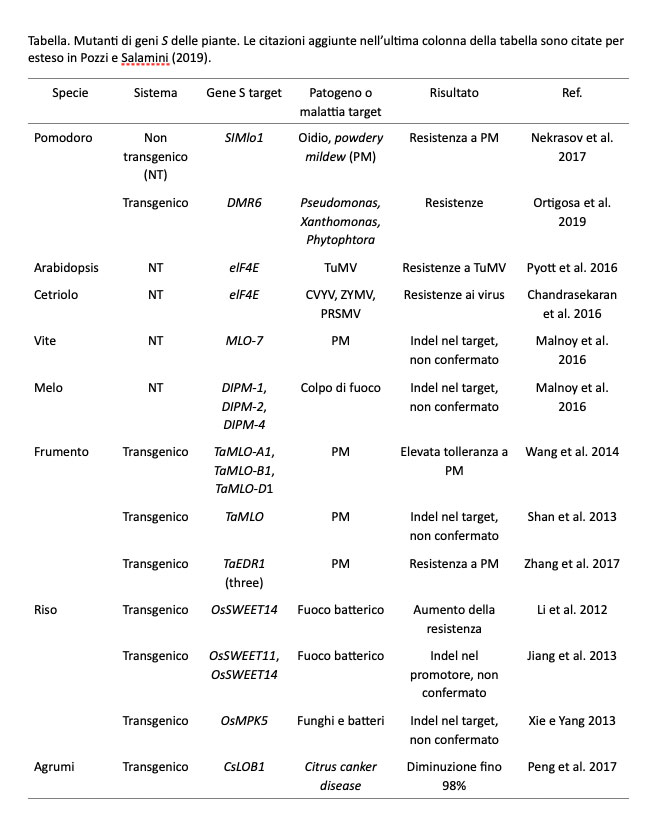
Le resistenze recessive dei geni S sono associate a effetti pleoiotropici che riducono la fitness della pianta. Una strategia per risolvere questo problema è di utilizzare la variabilità presente nel germoplasma di una specie, associandola a sequenziamenti e fenotipizzazioni per resistenza ed effetto pleiotropico, con l’obiettivo di reperire alleli S deboli. Alternativamente, ci si può concentrare sulla individuazione precisa dei meccanismi molecolari della LOS alla ricerca di soppressori genetici o sull’analisi delle interazioni proteiche interessate. In senso applicativo è stata utilizzata, come è il caso dell’orzo, la selezione fenotipca contro gli effetti pleiotropici. In conclusione, il ricorso a condizioni di silenziamento di geni S ha aperto una nuova e sicura via alla produzione di piante immuni da malattie.
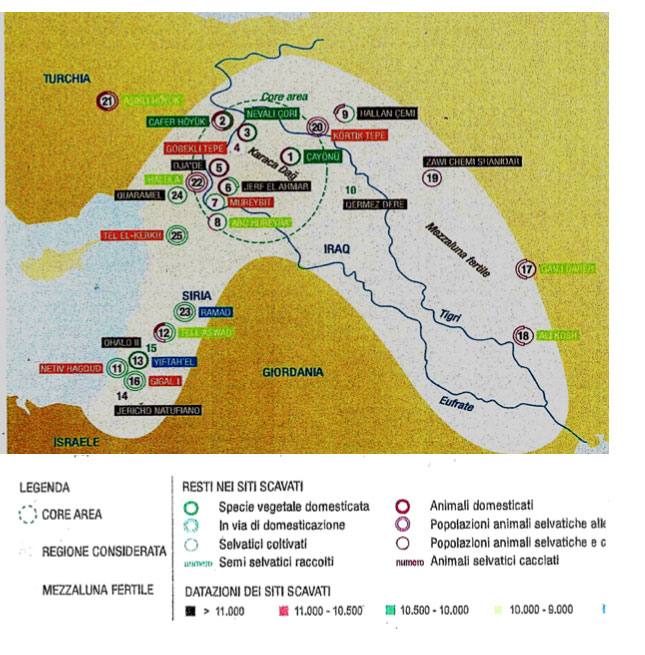
L’addomesticamento neolitico delle piante nell’Oriente Vicino. Quanto segue è ripreso e rielaborato da Salamini (2014). Circa 10.000 anni prima dell’inizio dell’agricoltura, nel Medio Oriente la raccolta sistematica a fini alimentari dei semi di piante selvatiche era già praticata: a Ohalo II, località a sud del corridoio palestinese, si raccoglievano ghiande, mandorle e semi dei progenitori selvatici del frumento monococco e dell’orzo. L’inizio dell’agricoltura del Vicino Oriente è riconducibile alla fine del Younger Dryas, un periodo di circa 1200 anni freddo e secco. Nell’area, l’addomesticamento dei cereali non è avvenuto prima di 11.000 anni fa, epoca corrispondente al periodo Neolitico pre-ceramico A (PPNA). Nel Neolitico pre-ceramico B (PPNB), nel sud-est dell’Anatolia compaiono forme addomesticate. L’orzo coltivato, addomesticato nella valle del Giordano (Badr et al., 2002), si ritrova come tale a metà del PPNB. Le piante agrarie addomesticate nel Medio Oriente sono frumento diploide (monococco), orzo, frumento tetraploide (dicocco), lenticchia, pisello, cece, veccia amara, lino, segale. Tipizzando il DNA di un gruppo di linee selvatiche di monococco, a noi fu possibile notare che le linee con il DNA più simile a quello delle varietà coltivate provenivano dalle alture del Karaca Dagˇ in Turchia, poste tra i corsi superiori del Tigri e dell’Eufrate, nella parte centrale della cosiddetta Mezzaluna Fertile (Heun et al., 1997). Il monococco è probabilmente il primo cereale addomesticato, quindi la sua patria poteva aver assistito all’emergere dell’agricoltura. Dalla stessa regione della Turchia sud-orientale provengono i progenitori selvatici dei frumenti tetraploidi coltivati: grano duro e farro (Salamini et al., 2002; Ozkan et al., 2002; 2005). Negli strati scavati corrispondenti al Neolitico preceramico B di villaggi agricoli di questa regione, come Çafer Höyük, Çayönü e Nevali Çori, sono stati ritrovati i semi carbonizzati dei primi frumenti addomesticati. Notando che il massiccio del Karaca Dagˇ è incluso nella distribuzione geografica dei progenitori selvatici di diverse piante coltivate, nel 2000, su «Science», un gruppo di scienziati israeliani (Lev-Yadun, 2000) concluse che in questa regione (core area) si era sviluppata l’innovazione agricola che negli anni sessanta del secolo scorso l’archeologo australiano Vere Gordon Childe per la prima volta definì «rivoluzione neolitica»: invenzione dell’agricoltura, stile di vita sedentario, stratificazione sociale, riti civili e sacerdotali. Childe (1951) si sbagliava, però, quando considerava l’alto Tigri ed Eufrate una neoliticizzazione secondaria. Klaus Schmidt (2011) segnala che negli stessi anni Robert e Linda Braidwood suggerivano che la core area di sviluppo agricolo era da individuare nelle colline pedemontane delle catene montuose del Tauro e degli Zagros (Zender, 2011). Successivi studi sulla geografia dell’addomesticamento del monococco, basati sulla sequenza del DNA di 18 geni (Kilian et al., 2007), confermarono che il progenitore selvatico di questa specie (razza β di Triticum boeoticum) proviene dalle alture del Karaca Dag˘, al centro della core area; la razza β di questa specie selvatica è probabilmente stata coltivata tal quale, dando poi origine a domesticazioni indipendenti. Questo «modello di dispersione specifica» deriva da dati molecolari attestanti che la variabilità genetica nel gruppo di genotipi addomesticati è simile a quella dei selvatici, una constatazione che suggerisce l’estrazione successiva e ripetuta dalla razza β di varietà da coltivare. Anche grano duro e spelta sono derivati dalle popolazioni selvatiche di Triticum dicoccoides che ancora si trovano attorno alla catena montuosa del Karaca Dag˘. Le forme coltivate di frumento tetraploide potrebbero aver avuto, nel corso della domesticazione o dopo, ibridazioni con genotipi selvatici provenienti dalle montagne turche del Kartal-Kara Dag˘ e dall’Iran-Iraq. L’addomesticamento dell’orzo è avvenuto nel Levante del sud (Badr et al., 2000), come peraltro sostenuto dalla lunga storia d’uso dell’orzo selvatico fino dall’epipaleolitico del Vicino Oriente. La figura che segue riassume quanto presentato relativamente all’addomesticamento delle piante agrarie e degli animali nel Vicino Oriente.
Altre linee di ricerca sviluppate dallo scrivente sono elencate nella Nota 2 che riporta anche le relative indicazioni bibliografiche.
Nota 1. CV. Nato nel marzo 1939. Laureato nel 1963 in Scienze Agrarie. Libero docente in Genetica Vegetale nel 1971. Dal 1966 al 1975 Ricercatore negli Istituti del Ministero dell’Agricoltura. Dal 1969 al 1970 Ricercatore associato, Purdue University, USA. Dal 1971 al 1974 Professore di Botanica all’Università di Piacenza. Nel 1977 Ricercatore, Brookeven National Laboratory, USA. Dal 1985 al 2004 Direttore, Dipartimento di Miglioramento Genetico e Fisiologia delle Piante, Colonia, Max-Planck-Institut für Züchtungsforschung. Nello stesso Istituto svolge la funzione di Direttore generale per due periodi di 3 anni. Dal 1990 a tuttora Professore onorario, Università di Colonia. Dal 1997 al 2009 (anno del congedo) Professore ordinario, Università di Potenza, Verona, Milano. Nel 2004 riceve la Laurea Honoris Causa dall’Università di Bologna. È stato Direttore di Istituti italiani e stranieri e Coordinatore di programmi di ricerca nazionali. Dal 1994 al 1996 ha svolto la funzione di Presidente della Società Italiana di Genetica Agraria. Ha accettato incarichi di consulenza scientifica in Italia, Germania, Francia ed è stato membro di Comitati scientifici presso la Fondazione Parco Tecnologico Padano, Lodi, CIVR, comitato Scienze agrarie e Scienze veterinarie e Scienze e Tecnologie degli alimenti, Expo 2015, Milano e Coordinatore del Comitato MIUR di selezione presidenti e membri del CDA degli Enti Nazionali di Ricerca. È membro affiliato alla Max-Planck-Gesellschaft, all’Accademia Nazionale dei Lincei, all’EMBO, all’Accademia Europaea, all’Accademia dei Georgofili e all’Accademia Nazionale di Agricoltura. La sua attività scientifica è stata riconosciuta con la Medaglia dei XL, il premio alla carriera della Fondazione Invernizzi, la commemorazione delle riviste Maidica e Plant Breeding Reviews, i premi Scanno (alimentazione) e Galileo Galilei. Ha costituito 6 novità vegetali e ottenuto 10 brevetti biotecnologici; le pubblicazioni sono oltre 600 e 10 le presenze in Comitati editoriali.
Nota 2. Ricerche svolte e relativa bibliografia. I mutanti glossy di mais (seguito della tesi di laurea): Salamini F., C. Lorenzoni, 1970. Genetical analysis of glossy mutants of maize. III. Intracistron recombination and high negative interference at the gl1 locus. Mol. Gen. Genet. 105:225-232. Bianchi G., P. Avato, F. Salamini, 1978. Glossy mutants of maize. VIII. Accumulation of fatty aldehydes in surface waxes of gl5 maize seedlings. Biochemical Genetics 16:1015-1021. Carboidrati nel mais: Tsai, C.Y., F. Salamini, O.E. Nelson, 1970. Enzymes of carbohydrate metabolism in the developing endosperm of maize. Plant Physiol. 46:299-306. Gentinetta, E., M. Zambello, F. Salamini, 1979. Free sugars in developing maize grain. Cereal Chem. 56:81-83.Le orticole a Montanaso: Baldi G., F. Salamini, 1973. Variability of essential amino acid content in seeds of 22 Phaseolus species. Theor. Appl. Genet. 43:75-78. Soressi G.P., E. Gentinetta, M. Odoardi, F. Salamini, 1974. Leaf peroxidase activity in tomato mutants affecting plant morphology. Biochem. Genetics 12:181-198. Zeine e Opaco2: Giannazza E. et al., 1977. Amino acid composition of zein molecular components. Phytochemistry 15:315-317. Soave C., R. Reggiani, N. Di Fonzo, F. Salamini,1981. Clustering of genes for 20 kd zein subunits in the short arm of maize chromosome 7. Genetics 97:363-377. Motto M., et al., 1988. Molecular cloning of the o2-m5 allele of Zea mays using transposon marking. Mol. Gen. Genet. 212:488-494. Hartings H., et al., 1989. The O2 gene which regulates zein deposition in maize endosperm encodes a protein with structural homologies to transcriptional activators. EMBO J. 8:2795-2801. Lohmer S., et al., 1991. The maize regulatory locus Opaque-2 encodes a DNA-binding protein which activates the transcription of the b-32 gene. EMBO J. 10:617-624. Lohmer S., et al., 1993 Translation of the mRNA of the maize transcriptional activator Opaque-2 is inhibited by upstream open reading frames present in the leader sequence. Plant Cell 5:65-73. Elementi trasponibili: Salamini F., 1981. Controlling elements at the Opaque-2 locus of maize: their involvement in the origin of spontaneous mutation. Cold Spring Harbor Symp. Quant. Biol. 45:467-476. Döring H.P., M. Freeling, M.A. Johns, P. Kunze, A. Merckelbach, F. Salamini, and P. Starlinger, 1984. A Ds-mutation of the Adh1 gene in Zea mays L. Mol. Gen. Genet. 193:199-204. Hartings H., et al., 1991. Nucleotide sequence of the Bg transposable element of Zea mays L. Maydica 36:355-359. Mappe molecolari: Gebhardt C., et al., 1989. RFLP analysis and linkage mapping in Solanum tuberosum. Theor. Appl. Genet. 78:65-75. Gebhardt C., et al., 1991. RFLP maps of potato and their alignment with the homoeologous tomato genome. Theor. App. Genet. 83:49-57. Barzen E., W. Mechelke, E. Ritter, J.F. Seitzer, F. Salamini, 1992. RFLP markers for sugar beet breeding: chromosomal linkage maps and location of major genes for rhizomania resistance, monogermy and hypocotyl colour. Plant J. 2:601-611. Taenzler B., et al., 2002. Molecular linkage map of Einkorn wheat: mapping of storage –protein and soft-glume genes and bread-making quality QTLs. Genetical Res. Camb. 80:131-143. Le piante resurrection: Bartels D., K. Schneider, Terstappen, G., Piatkowski, D., F. Salamini, 1990. Molecular cloning of abscisic acid-modulated genes which are induced during desiccation of the resurrection plant Craterostigma plantagineum. Planta 181:27-34. Bernacchia G. et al.,1995. The transketolase gene family of the resurrection plant Craterostigma plantagineum: differential expression during rehydration phase. EMBO J. 14:610-618. Bartels D., F. Salamini, 2001. Desiccation tolerance in the resurrection plant Craterostigma plantagineum: a contribution to the study of drought tolerance at the molecular level. Plant Physiol. 127:1346-1353. Orzo: Müller K.J., et al., 1995. The barley Hooded mutation caused by a duplication in a homeobox gene intron. Nature 374:727-730. Badr A., K. et al., 2000. On the origin and domestication history of barley (Hordeum vulgare). Molecular Biol. Evol. 17:499-510. Geni per la resistenza alle malattie: Leister D., A. Ballvora, F. Salamini, C. Gebhardt, 1996. A PCR-based approach for isolating pathogen resistance genes from potato with potential for wide application in plants. Nature Genetics 14:421-429. Büschges R., et al., 1997. The barley Mlo gene: a novel control element of plant pathogen resistance. Cell 88:695-705. Ballvora A. et al., 2002. The R1 gene for potato resistance to late blight (Phytophthora infestans) belongs to the leucine zipper/NBS/LRR class of plant resistance genes. Plant J. 30:361-371. Paal J., H. et al., 2004. Molecular cloning of the potato Gro 1-4 gene conferring resistance to pathotype Ro 1 of the root cyst nematode Globodera rostochensis, based on a candidate gene approach. Plant J. 38:285-297. L’origine delle piante agrarie: Heun M., R. Schäfer-Pregl, D. Klawan, R. Castagna, M. Accerbi, B. Borghi, F. Salamini, 1997. Site of Einkorn wheat domestication identified by DNA fingerprinting. Science 278:1312-1314. Salamini F., 1999. Il frumento monococco e l’origine dell’agricoltura. Le Scienze 373:68-74. Salamini F., H. Özkan, A. Brandolini, R. Schäfer-Pregl, W. Martin, 2002. Genetics and geography of wild cereal domestication in the Near East. Nature Reviews Genetics 3:429-441.Salamini F., 2003. Hormones and the green revolution. Science 302:71-72. Fotosintesi: Varotto C., et al., 2000. Disruption of the Arabidopsis photosystem I gene psaE1 affects photosynthesis and impairs growth. Plant J. 22:115-124. Kurth J., et al., 2002. Gene-sequence-tag expression analyses of 1,800 genes related to chloroplast functions. Planta 215:101-109. Richly E., et al., 2003. Co-variations in the nuclear chloroplast transcriptome reveal a regulatory master switch. EMBO Reports 4:491-498. Genomi: Velasco R, et al., 2007. A high quality draft consensus sequence of the genome of a heterozygous grapevine variety. PLoS ONE, Dec 2007, Issue 12 e1326. Velasco R, et al., 2010. The genome of the domesticated apple (Malus xdomestica Borkh). Nat. Genet. 42: 833-839. Verde I., et al., 2013. The high-quality draft genome of peach (Prunus persica) identifies unique patterns of genetic diversity, domestication and genome evolution. Nature Genetics. doi:10.1038/ng.2586.
Bibliografia
Badr A., Müller K., Schäfer-Pregl R., El Rabey H., Effgen S., Ibrahim H. H., Pozzi C., Rohde W., F. Salamini, 2000. On the origin and domestication history of barley (Hordeum vulgare). Molecular Biol. Evol. 17: 499-510.
Büschges R., Hollricher K., Panstruga R., Simons G., Wolter M., Frijters A., van Daelen R., van der Lee T., Diergaarde P., Groenendijk J., Töpsch S., Vos P., Salamini, F., P. Schulze-Lefert, 1997 The barley Mlo gene: a novel control element of plant pathogen resistance. Cell 88: 695-705.
Childe V. G., 1951. Man makes himself. New American library of world literature, New York.
Gavazzi G., F. Salamini, 1980. Mutagenesi e miglioramento delle piante coltivate. Quaderni di Biologia, L. De Carli (Ed). Piccin, Padova.
Hinze K., Thompson R.D., Ritter E., Salamini F., P. Schulze-Lefert, 1991. RFLP-mediated targeting of the mlo resistance locus in barley (Hordeum vulgare). Proc. Natl. Acad. Sci. USA 88: 3691-3695.
Lev-Yadun S., 2000. The cradle of agriculture. Science 288: 1602-1603.
Malnoy M., Viola R., Jung M.H., Koo O.J., Kim S., Kim J.S., Velasco R., C. N. Kanchiswamy, 2016. DNA-free genetically edited grapevine and apple protoplast using CRISPR/Cas9 ribonucleoproteins. Frontiers in Plant Science 7 doi:ARTN 1904 10.3389/fpls.2016.01904
Ozkan H., Brandolini A., Pozzi C., Effgen S., Wunder J., F. Salamini, 2005. A reconsideration of the domestication geography of tetraploid wheats. Theor Appl Genet 110: 1052-1060.
Özkan H., Brandolini A., Schäfer-Pregl R., F. Salamini, 2002. AFLP analysis of a collection of tetraploid wheats indicates the origin of emmer and hard wheat domestication in southeast Turkey. Mol. Biol. Evolution 19: 1797-1801.
Kilian B., Ozkan H., Walther A., Kohl J., Dagan T., Salamini F., W. Martin, 2007. Molecular diversity at 18 loci in 321 wild and 92 domesticated lines reveal no reduction of nucleotide diversity during Triticum monococcum (Einkorn) domestication: implications for the origin of agriculture. Mol Biol Evol. 24: 2657–2668.
Pessina S., Lenzi L., Perazzolli M., Campa M., Dalla Costa L., Urso S., Valè G., Salamini F., Velasco R., Malnoy M., 2016. Knockdown of MLO genes reduces susceptibility to powdery mildew in grapevine. Horticulture Research 3, no.16016.
Pozzi C., F. Salamini, 2019. Riassetto dei sistemi agricoli, priorità ad ecologia e sostenibilità. Rivista di Frutticoltura 83: 22-33.
Salamini F., 2014. Alle origini dell’Eden. Le Scienze: 63- 69.
Salamini F., Özkan H., Brandolini A., Schäfer-Pregl R., W. Martin, 2002. Genetics and geography of wild cereal domestication in the Near East. Nature Reviews Genetics 3: 429-441.
Schmidt K., 2011. Costruirono i primi templi. Oltre Edizioni, Genova.
Simons G., van der Lee T., Diergaarde P., van Daelen R., Groenendijk J., Frijters A., Büschges R., Hollricher K., Töpsch S., Schulze-Lefert P., Salamini F., Zabeau F., P. Vos, 1997. AFLP-based fine mapping of the Mlo gene to a 30 kb DNA segment of the barley genome. Genomics 44: 61-70.
Wolter M., Hollricher K., Salamini F., P. Schulze-Lefert, 1993. The mlo resistance alleles in barley trigger a defense mimic phenotype. Mol. Gen. Genet. 239: 122-128.
Zaidi S.S., Mukhtar M.S., S. Mansoor, 2018. Genome Editing: targeting susceptibility genes for plant disease resistance. Trends Biotechnol 36:898-906.
Zender M. A., 2011. The origins of agricolture in the Near East. Current Anthropology 52: 221-235.